Ruoansulatus
Kokeneet kirjoittajat eivät ole vielä tarkistaneet sivun nykyistä versiota, ja se voi poiketa merkittävästi 17. joulukuuta 2020 tarkistetusta versiosta . vahvistus vaatii 31 muokkausta .Ruoansulatus - kemiallinen (pääasiassa entsymaattinen), joskus myös mekaaninen ruoan prosessointi - joukko prosesseja, jotka varmistavat ravinteiden hajoamisen komponenteiksi, jotka ovat sopivia imeytymiseen ja aineenvaihduntaan osallistumiseen [1] . Ruoansulatuksen aikana ruoan orgaaniset makromolekyylit muuttuvat pienemmiksi molekyyleiksi, erityisesti elintarvikebiopolymeerien hajoaminen monomeereiksi . Tämä prosessi suoritetaan ruoansulatusentsyymien (hydrolyyttisten) entsyymien avulla . Yllä olevan käsittelyprosessin jälkeen ruoka imeytyy suoraan solujen sytoplasmaan (bakteerien ja sienten solunulkoisen ruoansulatuksen ja solunsisäisen ruoansulatuksen aikana) tai ruoansulatuskanavan seinämien kautta kehon nesteisiin (ihmisillä vereen tai imusolmukkeisiin).
Ruoansulatuksen biologinen merkitys
- Suurten hiukkasten pilkkominen pienemmiksi on välttämätöntä ruoan imeytymiseksi - sen kuljettaminen solujen sytoplasmaan solukalvon kautta ja eläimillä, joilla on suolensisäinen ruoansulatus - imeytyminen maha-suolikanavan seinämien läpi kuljetusjärjestelmään ( veri, imusolmukkeet ja niin edelleen).
- Pilkkominen proteiinien, DNA:n (ja osittain myös muiden elintarvikepolymeerien) monomeereiksi on välttämätöntä tämän tyyppisille organismeille spesifisten "omien" biomolekyylien myöhempää synteesiä varten monomeereistä.
Tärkeimmät ruuansulatuksen tyypit ja niiden jakautuminen elävien organismien kesken
Solunulkoinen (kaukainen) ruoansulatus
Tämäntyyppiselle ruoansulatukselle on ominaista se, että erityssolujen syntetisoimat entsyymit vapautuvat solunulkoiseen ympäristöön, jossa niiden hydrolyyttinen vaikutus toteutuu. Hydrolyyttisten entsyymien erittyminen soluista ruuansulatusonteloihin ilmestyi ensin koelenteraateissa ja ktenoforeissa . Annelideilla , äyriäisillä , hyönteisillä , pääjalkaisilla , vaippaeläimillä ja keuhkoeläimillä ( lukuun ottamatta lansettia ) solunulkoinen ruoansulatus on vallitseva. Se on kehitetty erityisesti hyvin järjestäytyneille eläimille ja ihmisille, joissa se tarjoaa ravintoaineiden alkusulatuksen. Näissä organismeissa erityssolut sijaitsevat riittävän kaukana ruoansulatusonteloista, joissa hydrolaasien toiminta toteutuu , joten solunulkoinen ruoansulatus määritellään etäiseksi. Erikoisonteloissa tapahtuvaa kaukaista ruoansulatusta kutsutaan onkaloksi. [2]
Solunulkoinen ruuansulatus on ominaista kaikille heterotrofisille organismeille, joiden soluissa on soluseinä - bakteerit, arkeat, sienet, lihansyöjäkasvit jne. Tällä ruoansulatusmenetelmällä entsyymejä erittyvät ulkoiseen ympäristöön tai ne kiinnittyvät ulkokalvolle (grammoina). negatiiviset bakteerit) tai soluseinässä. Ruoan sulatus tapahtuu solun ulkopuolella, jolloin muodostuneet monomeerit imeytyvät solukalvon kuljettajaproteiinien avulla .
Kavitaarinen ruoansulatusKavitaarinen ruoansulatus on ominaista monisoluisille eläimille, joilla on maha-suolikanava , ja se tapahtuu jälkimmäisen ontelossa.
Ekstraintestinaalinen ruoansulatusSuolen ulkopuolinen ruoansulatus on tyypillistä joillekin eläimille, joilla on suolisto, mutta jotka ruiskuttavat ruoansulatusentsyymejä saaliin kehoon ja imevät sitten puolisulatettua ruokaa ( hämähäkit ja uimakuoriaisen toukat ovat tunnettuja esimerkkejä tällaisista eläimistä).
Kalvo (parietaalinen) ruoansulatusParietaalinen ruoansulatus suoritetaan limakerroksessa ohutsuolen mikrovillien välissä ja suoraan niiden pinnalla ( glykokalyyksissä ) selkärankaisilla ja joillakin muilla eläimillä.
Solunsisäinen ruoansulatus
Solunsisäinen digestio on prosessi, joka liittyy läheisesti endosytoosiin , ja se on ominaista vain niille eukaryoottiryhmille, joilta puuttuu soluseinä (jotkut protistit ja useimmat eläimet). Fylogeneettisesti tämäntyyppinen ruoansulatus on vanhin, koska se on yleistä yksinkertaisimmissa ja primitiivisimmissä monisoluisissa organismeissa, usein lattamatoissa . Solunsisäistä ruoansulatusta on kahta tyyppiä. Ensimmäinen liittyy pienten molekyylien kuljettamiseen solukalvojen läpi ja sitä seuraavaan pilkkomiseen sytosolientsyymeillä . Solunsisäinen ruoansulatus voi tapahtua myös erikoistuneissa solunsisäisissä onteloissa - ruoansulatusvakuoleissa, jotka ovat jatkuvasti läsnä tai muodostuvat fagosytoosin ja pinosytoosin aikana ja katoavat, kun siepatun ruoan pilkkoutuminen on päättynyt. Tämä toinen solunsisäinen digestiotyyppi liittyy useimmissa tapauksissa lysosomien osallistumiseen, jotka sisältävät laajan valikoiman erilaisia hydrolyyttisiä entsyymejä, joiden optimaalinen pH on 3,5-5,5, riippumatta siitä, mihin organismiin ne kuuluvat. Lysosomientsyymit eroavat merkittävästi solunulkoiseen ruoansulatukseen osallistuvista. [2]
Perisellulaarisessa ympäristössä olevat ruokahiukkaset tai ruokaliuokset aiheuttavat plasmakalvon invaginaatioita, jotka sitten kiinnittyvät ja uppoavat sytoplasmaan muodostaen pino- ja fagosyyttisiä vakuoleja. Yhdistämällä jälkimmäiseen lysosomit muodostavat niin sanottuja fagosomeja , joissa tapahtuu entsyymien kosketus vastaavien substraattien kanssa. Tällöin tapahtuu solunsisäistä ekstraplasmista digestiota, jonka fysikaalis-kemialliset lait eivät eroa tai eroavat vähän vatsansulatuksen laeista. Tästä syystä toisen tyypin solunsisäistä hajoamista voidaan luonnehtia mikroonteloksi. Koska lysosomien ja vakuolien kalvon sisäpinta sisältää erilaisia hydrolyyttisiä entsyymejä, voidaan mikroontelohydrolyysiä täydentää kalvohydrolyysillä. Syntyneet hydrolyysituotteet imeytyvät fagosomikalvon läpi. Ruoansulatussyklin päätyttyä fagosomijäännökset työntyvät ulos solun ulkopuolelle eksosytoosin avulla . Lysosomeilla on tärkeä rooli myös solun omien rakenteiden hajoamisessa, joita käytetään ravintoaineena joko solussa tai sen ulkopuolella. [2]
Ruoansulatus eläimissä
Useimmissa eläimissä suolensisäinen ruoansulatus yhdistetään solunsisäiseen ruoansulatukseen. Sienissä on vain solunsisäistä ruoansulatusta . Hyönteisille , sukkulamadoille ja selkärankaisille on ominaista pääasiassa suolensisäinen ruuansulatus (joskus täydennettynä suoliston ulkopuolisella) . Joillakin eläimillä suolet puuttuvat (sienet, placozoa, cnidosporidia, dicyemids ja orthonektidit, pogonoforit, heisimadot, trematodien sporokystit, rhizocephalic rapujen aikuiset vaiheet ja tietyt muiden ryhmien lajit). Tällaiset eläimet käyttävät solunsisäistä ruoansulatusta (sienet, placozoans), suoliston ulkopuolista ulkoista tai parietaalista ruoansulatusta (placozoans, heisimadot) tai ruokkivat endosymbionteja. jotka voidaan sulattaa solunsisäisesti (pogonoforit, jotkut simpukat).
Joillakin eläimillä (cnidarians, nemertodermatids ja xenoturbellids, monet flatmadot) suolet ovat sokeasti suljettuina (vain suuaukko johtaa siihen). Samaan aikaan kehittyy usein lihaksikas nielu, johon turbellareissa nielurauhaset avautuvat vapauttaen ruoansulatusentsyymejä.
Useimmilla eläimillä ruoansulatuskanava on läpikäynyt. Se sisältää etu-, keski- ja takasuolen; etu- ja takasuoli kehittyvät ektodermista, keskisuoli endodermista. Ruoansulatuskanava on usein jaettu toiminnallisiin osiin - suuonteloon, nieluun, ruokatorveen, mahalaukkuun ja suolistoon. Nilviäisten ja useimpien niveljalkaisten ruuansulatukseen osallistuvat ruoansulatusrauhaset - hepatohaima, jonka kanavat avautuvat mahalaukkuun, ja usein myös sylkirauhaset.
Niveljalkaisissa etusuolassa (erityisesti mahassa) joissakin pääasiassa kasvinsyöjälajeissa on kitiiinimuodostelmia, jotka toimivat kiinteän ruoan jauhamiseen. Suun laite muodostuu muunnetuista raajoista.
Ruoansulatus selkärankaisilla on yhdistelmä seuraavista toisiinsa liittyvistä prosesseista: ruuan mekaaninen ja fyysinen prosessointi, sen liikkuminen ruoansulatuskanavan läpi, ruoan komponenttien kemiallinen tuhoutuminen ( hydrolyysi ), joka tapahtuu maha-suolikanavan eritystoiminnolla ; orgaanisten ja epäorgaanisten yhdisteiden, mukaan lukien hivenaineet ja vesi, imeytymisprosessi vereen ja imusolmukkeisiin ; kehon poistettavien kuona-aineiden erittyminen maha-suolikanavan onteloon ; niiden poistuminen kehosta yhdessä sulamattomien ruokajäämien kanssa.
Selkärankaisille on ominaista solunsisäisen ruuansulatuksen puuttuminen tai heikko ilmentyminen sekä suolensisäisen ja parietaalisen ruoansulatuksen vallitsevuus. Kaikilla selkärankaisilla maksa ja haima osallistuvat ruoansulatukseen, maaeläimillä myös sylkirauhaset.
Ihmisen ruoansulatusprosessi
Suuontelo
Ihmisellä ruoansulatus alkaa suussa , jossa ruoka pureskellaan. Tämä prosessi stimuloi eksokriiniset rauhaset erittämään sylkeä . Syljessä oleva amylaasi osallistuu polysakkaridien hajoamiseen ja boluksen - ruokapalan - muodostumiseen, mikä helpottaa ruoan kulkeutumista ruokatorven läpi . Nielun limakalvon reseptorien ärsytys aiheuttaa nielemisrefleksin , joka koordinoituu nielemiskeskuksessa, joka sijaitsee ytimessä ja pons varoliissa . Koordinoitu nieleminen sisältää pehmeän kitalaen ja uvulan , jotka estävät ruokaa pääsemästä nenäonteloon , sekä kurkunpään , joka estää ruoan pääsyn henkitorveen .
Vatsa
Vatsa sijaitsee pallean alla vasemmassa hypokondriumissa ja epigastrisessa alueella. On 3 skiniä:
- Ulkoinen (vatsakalvo)
- lihaskerros
- ulkokerros (pitkittäinen)
- keskikerros (pyöreä);
- sisäkerros (vino).
- Sisäinen (limakalvo) - vuorattu keratinisoitumattomalla, lieriömäisellä epiteelillä.
Ruoka tulee vatsaan kulkemalla sydämen sulkijalihaksen läpi . Siellä se sekoittuu mahanesteeseen , jonka aktiiviset komponentit ovat suolahappo ja ruoansulatusentsyymit :
- Pepsiini - hajottaa proteiinit aminohapoiksi , polypeptideiksi , oligopeptideiksi [3] .
- Renniini (tai kymosiini ) - saatavilla alle 1-vuotiaille lapsille, auttaa sulattamaan maitotuotteita . Vuoden kuluttua kymosiini katoaa, sen toiminnot suorittaa kloorivetyhappo.
Vatsan parietaalisolut erittävät myös Castlen luontaista tekijää , joka on välttämätön B 12 -vitamiinin imeytymiselle .
Ohutsuoli
Pylorisen sulkijalihaksen kautta ruoka tulee ohutsuoleen . Ohutsuolen ensimmäinen osa on pohjukaissuoli , jossa ruoka sekoitetaan sappeen , joka emulgoi rasvoja haiman ja ohutsuolen entsyymeillä, jotka hajottavat hiilihydraatteja ( maltoosi , laktoosi , sakkaroosi ), proteiineja ( trypsiini ja kymotrypsiini ). Ohutsuolessa pääasiallinen määrä ravintoaineita ja vitamiineja imeytyy suolen seinämän läpi.
Paksusuoli
Ohutsuolen läpi kulkemisen jälkeen ruoka menee paksusuoleen , joka koostuu umpisuolesta , paksusuolesta , sigmoidisesta paksusuolesta ja peräsuolesta . Täällä tapahtuu veden ja elektrolyyttien imeytyminen, ja täällä tapahtuu myös ulosteiden muodostumista .
Ruoansulatuksen säätely
Ruoansulatus on ihmisellä psykofysiologinen prosessi. Tämä tarkoittaa, että reaktioiden järjestykseen ja nopeuteen vaikuttavat maha-suolikanavan humoraaliset kyvyt , ruoan laatu ja autonomisen hermoston tila .
Ruoansulatukseen vaikuttavat humoraaliset kyvyt määräytyvät mahalaukun ja ohutsuolen limakalvojen solujen tuottamien hormonien avulla. Tärkeimmät ruoansulatushormonit ovat gastriini , sekretiini ja kolekystokiniini . Ne vapautuvat maha-suolikanavan verenkiertojärjestelmään ja edistävät ruoansulatusnesteiden tuotantoa ja ruoan liikkumista.
Ruoan sulavuus riippuu ruuan laadusta [4] :
- huomattava kuitupitoisuus (mukaan lukien liukoinen) voi merkittävästi vähentää imeytymistä;
- jotkin ruoan sisältämät hivenaineet vaikuttavat aineiden imeytymiseen ohutsuolessa [5] ;
- erilaiset rasvat imeytyvät eri tavoin. Tyydyttyneet eläinrasvat imeytyvät ja muuttuvat ihmisrasvoiksi paljon helpommin kuin monityydyttymättömät kasvirasvat, jotka eivät käytännössä osallistu ihmisrasvan muodostumiseen;
- hiilihydraattien, rasvojen ja proteiinien imeytyminen suolistosta vaihtelee jonkin verran vuorokaudenajan ja vuodenajan mukaan;
- imeytyminen vaihtelee myös sen mukaan, mikä on aikaisemmin suolistossa joutuneiden tuotteiden kemiallinen koostumus.
Ruoansulatusta säätelee myös autonominen hermosto . Parasympaattinen osa stimuloi eritystä ja peristaltiikkaa , kun taas sympaattinen osa estää .
Ruoansulatukseen vaikuttavat hormonit ja muut biologisesti aktiiviset aineet
Gastroenteropankreaattinen endokriininen järjestelmä - endokriinisen järjestelmän osasto , jota edustavat endokriiniset solut ( apudosyytit ) ja peptidergiset neuronit , jotka tuottavat peptidihormoneja hajallaan ruoansulatusjärjestelmän eri elimiin . Se on tutkituin osa diffuusista endokriinistä järjestelmää (synonyymi APUD-järjestelmälle ) ja sisältää noin puolet sen soluista. Gastroenteropankreaattista endokriinistä järjestelmää on kutsuttu "ihmiskehon suurimmaksi ja monimutkaisimmaksi endokriiniseksi elimeksi" [6] .
Gastriini on mahalaukun G-solujen syntetisoima hormoni, joka sijaitsee pääasiassa mahalaukun pylorisessa alueella. Gastriini sitoutuu tiettyihinmahalaukun gastriinireseptoreihin . Lisääntynyt adenylaattisyklaasiaktiivisuusmahalaukun parietaalisoluissa lisää mahanesteen , erityisesti suolahapon, eritystä . Gastriini lisää myös mahalaukun pääsolujen pepsiinin eritystä , mikä yhdessä mahanesteen happamuuden lisääntymisen kanssa tarjoaa optimaalisen pH :n pepsiinin toiminnalle ja edistää ruoan optimaalista sulamista mahalaukussa. Samaan aikaan gastriini lisää bikarbonaattien ja liman eritystä mahalaukun limakalvossa ja suojaa siten limakalvoa suolahapon ja pepsiinin vaikutuksilta. Gastriini estää mahalaukun tyhjenemistä, mikä varmistaa riittävän pitkäkestoisen suolahapon ja pepsiinin altistumisen ruokabolukselle ruoansulatusta varten. Lisäksi gastriini lisää prostaglandiini E:n tuotantoa mahalaukun limakalvossa, mikä johtaa paikalliseen vasodilataatioon, lisääntyneeseen verenkiertoon ja mahalaukun limakalvon fysiologiseen turvotukseen sekä leukosyyttien kulkeutumiseen mahan limakalvolle.
Sekretiini on peptidihormoni , joka koostuu 27 aminohappotähteestä , jota tuottavat ohutsuolen limakalvon S-solut ja joka osallistuu haiman eritystoiminnan säätelyyn. Sappihapot lisäävät sekretiinin tuotannon stimulaatiota [6] . Vereen imeytyessään sekretiini saavuttaa haiman, jossa se lisää veden ja elektrolyyttien, pääasiassa bikarbonaatin , eritystä . Lisäämällä haiman erittämän mehun määrää sekretiini ei vaikuta rauhasen entsyymien muodostumiseen. Tämän toiminnon suorittaa toinen ohutsuolen limakalvossa tuotettu aine - kolekystokiniini . Sekretiinin biologinen määritelmä perustuu sen kykyyn (annettaessa eläimille suonensisäisesti) lisätä alkalin määrää haimanesteessä [7] . Sekretiini estää suolahapon tuotantoa mahalaukun parietaalisoluissa [8] . Pääasiallinen sekretiinin aiheuttama vaikutus on bikarbonaattien tuotannon stimulaatio sappiepiteelin , haimatiehyiden ja Brunnerin rauhasten toimesta , mikä tuottaa jopa 80 % bikarbonaattien erityksestä vastauksena ruoan nauttimiseen. Tämä vaikutus välittyy kolekystokiniinin erittymisen kautta, ja tämä johtaa sapen tuotannon lisääntymiseen, sappirakon ja suoliston supistusten stimulaatioon ja suoliston mehun erittymisen lisääntymiseen [6] .
Kolekystokiniini ( CCK ; entinen pankreotsymiini ) on neuropeptidihormoni , jota tuottavat pohjukaissuolen limakalvon ja proksimaalisen tyhjäsuolen I-solut [9] . Kolekystokiniini toimii välittäjänä erilaisissa kehossa tapahtuvissa prosesseissa, mukaan lukien ruoansulatus. Kolekystokiniini stimuloi Oddin sulkijalihaksen rentoutumista ; lisää maksan sapen virtausta; lisää haiman eritystä; vähentää painetta sappijärjestelmässä: aiheuttaa pyloruksen supistumista , mikä estää sulatetun ruoan liikkumisen pohjukaissuoleen [10] [6] . Kolekystokiniini onmahalaukun parietaalisolujen suolahapon erityksen estäjä [8] . Somatostatiini on kolekystokiniinin estäjä.
Glukoosiriippuvainen insulinotrooppinen polypeptidi (aiemmin yleiset nimet: gastroinhibitorinen polypeptidi , mahaa estävä peptidi ; yleiset lyhenteet: GIP , GIP tai GIP ) on peptidihormoni , joka koostuu 42 aminohappotähteestä, joita tuottavat pohjukaissuolen ja proksimaalisen limakalvon K-solut jejunumin osa[ 6] . Kuuluu secretin -perheeseen . Glukoosiriippuvainen insulinotrooppinen polypeptidi on inkretiini , toisin sanoen sitä tuotetaan suolistossa vasteena suun kautta tapahtuvalle ruuan nauttimiselle. Glukoosiriippuvaisen insulinotrooppisen polypeptidin päätehtävä on stimuloida haiman beetasolujen insuliinin eritystä vasteena ravinnon saantiin. Lisäksi GIP estää rasvojen imeytymistä, estää natriumin ja veden takaisinimeytymistä ruoansulatuskanavassa , estää lipoproteiinilipaasia [6] .
Vasoaktiivinen intestinaalinen peptidi (kutsutaan myös vasoaktiiviseksi suolen polypeptidiksi ; yleiset lyhenteet VIP ja VIP ) on neuropeptidihormoni , joka koostuu 28 aminohappotähteestä, jota löytyy monista elimistä, mukaan lukien suolet , aivot ja selkäydin sekä haima [6] . Vasoaktiivinen suolen peptidi, toisin kuin muut sekretiiniperheen peptidihormonit, on yksinomaan välittäjäaine . Sillä on voimakas stimuloiva vaikutus suolen seinämän verenkiertoon sekä suolen sileisiin lihaksiin [6] . Se on estäjä, joka estää suolahapon erittymistä mahalaukun limakalvon parietaalisoluissa [ 11] . VIP on myös mahalaukun pääsolujen pepsinogeenituotannon stimulaattori[ 12 ] .
Motiliini on maha-suolikanavan , pääasiassa pohjukaissuolen ja tyhjäsuolen limakalvon kromafiinisolujen tuottama hormoni.
Somatostatiini on haiman Langerhansin saarekkeiden deltasolujen hormoni sekä yksi hypotalamuksen hormoneista . Kemiallisen rakenteensa mukaan se on peptidihormoni. Somatostatiini estää somatotropiinia vapauttavan hormonin eritystä hypotalamuksessa sekä somatotrooppisen hormonin ja kilpirauhasta stimuloivan hormonin eritystä aivolisäkkeen etuosassa . Lisäksi se myös estää erilaisten hormonaalisesti aktiivisten peptidien ja mahalaukussa , suolistossa , maksassa ja haimassa tuotetun serotoniinin erittymistä . Erityisesti se alentaa insuliinin , glukagonin , gastriinin , kolekystokiniinin , vasoaktiivisen suolen peptidin , insuliinin kaltaisen kasvutekijä-1 :n eritystä .
Ruoansulatusentsyymit
Ruoansulatusentsyymit ovat ryhmä entsyymejä , jotka hajottavat monimutkaiset ruoan komponentit kemiallisesti yksinkertaisemmiksi aineiksi, jotka sitten imeytyvät suoraan elimistöön tai pääsevät verenkiertoelimistöön . Laajemmassa merkityksessä ruoansulatusentsyymeillä tarkoitetaan myös kaikkia entsyymejä, jotka hajottavat suuria (yleensä polymeerisiä) molekyylejä monomeereiksi tai pienemmiksi osiksi. Ruoansulatusentsyymejä tuotetaan ja ne toimivat ihmisten ja eläinten ruoansulatusjärjestelmässä . Lisäksi lysosomien solunsisäiset entsyymit voidaan katsoa sellaisiksi entsyymeiksi . Ruoansulatusentsyymien pääasialliset toimintapaikat ihmisillä ja eläimillä ovat suuontelo , mahalaukku ja ohutsuole . Ruoansulatusentsyymejä tuottaa ruoansulatuselinten rauhaskudos: sylkirauhaset , maharauhaset, maksa , haima ja ohutsuolen rauhaset. Lisäksi suoliston pakollinen mikrofloora suorittaa osan entsymaattisista toiminnoista.
Suoliston mikrofloora
Ihmisen paksusuolessa elävät mikro-organismit erittävät ruoansulatusentsyymejä, jotka auttavat sulattamaan tietyntyyppisiä ruokia:
- E. coli - edistää laktoosin sulamista ;
- Laktobasillit - muuttavat laktoosia ja muita hiilihydraatteja maitohapoksi .
Hyönteissyöjäkasvien ruoansulatusentsyymit
Proteaasit eristettiin Nepenthes macferlanein erityksestä , ja myös lipaasiaktiivisuus osoitettiin. Sen pääentsyymi, nepentetsiini, muistuttaa pepsiiniä substraattispesifisyydessä [13] .
Katso myös
Muistiinpanot
- ↑ M. S. Giljarov. Biologinen tietosanakirja. – 1986.
- ↑ 1 2 3 A. M. Ugolev. Ruoansulatuksen evoluutio ja toimintojen kehittymisen periaatteet. – 1985.
- ↑ Jakubke H.-D., Eshkayt H. Aminohapot, peptidit, proteiinit. - M .: Mir, 1985. - 289 s.
- ↑ Faleev A. V. Harmonian taika Arkistokopio 23.6.2008 Wayback Machinessa - 2006. S. 6.
- ↑ Esimerkiksi L. Fakambin tieteelliset työt osoittivat, että kypsät juustot, joissa on paljon kalsiumia, säilyttävät osan rasvoista ja estävät niiden imeytymisen suolistosta. Näin ollen tämä rasvan osa ei pääse kehoon, vaan lähtee ulosteen mukana.
- ↑ 1 2 3 4 5 6 7 8 Maev I. V., Samsonov A. A. Pohjukaissuolen sairaudet. M. , MEDpress-inform, 2005, - 512 s, ISBN 5-98322-092-6 .
- ↑ Ruoansulatus // Suuri Neuvostoliiton Encyclopedia : [30 nidettä] / ch. toim. A. M. Prokhorov . - 3. painos - M . : Neuvostoliiton tietosanakirja, 1969-1978.
- ↑ 1 2 Butov M.A., Kuznetsov P.S. Ruoansulatuskanavan sairauksia sairastavien potilaiden tutkimus. Osa 1. Vatsasairauksista kärsivien potilaiden tutkimus Arkistoitu 22. syyskuuta 2011 Wayback Machinessa . Sisätautien propedeutiikan oppikirja lääketieteellisen tiedekunnan 3. vuoden opiskelijoille. Ryazan. 2007 (2,42 Mt).
- ↑ Murray R, Grenner D, Mays P, Rodwell W. Human Biochemistry Arkistoitu 23. tammikuuta 2015. . Osa 2, s. 272.
- ↑ Yakovenko E. P., Grigoriev P. Ya., Agafonova N. A., Yakovenko A. V. Kolereettisten lääkkeiden paikka kliinisessä käytännössä Arkistokopio päivätty 30. elokuuta 2009 Wayback Machinessa . Hoitava lääkäri. 2005, nro 6.
- ↑ Acid Conditions in Children Arkistoitu 8. marraskuuta 2006 Wayback Machinessa . Venäjän lääketieteen akatemian akateemikon, professori V. A. Tabolinin toimittamana . M., 1999, 120 s.
- ↑ Lyhyt G. F. Ruoansulatuskanavan fysiologia. - Krasnodar: 2009. - 608 s. Kustantaja OOO BK "Group B". ISBN 5-93730-021-1 .
- ↑ Zoltán A. Tökés, Wang Chee Woon ja Susan M. Chambers. Lihansyöjäkasvin Nepenthes macferlanei L. Planta, 1974, osa 119, numero 1, 39-46, erittämät ruoansulatusentsyymit
| |
| |||
|---|---|---|---|---|
| ||||
| ihmisen ruoansulatusjärjestelmä | ||
|---|---|---|
| Vatsan yläpuolella | 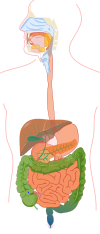 | |
| Vatsa | Pylorus | |
| Ohutsuoli | ||
| Kaksoispiste | ||
| peräaukko | ||
| Suuret ruuansulatusrauhaset |
| |
| Ruoansulatuksen fysiologia , ihmisen ruoansulatusjärjestelmä | |
|---|---|
| Enteraalinen hermosto | |
| Enterokriininen | |
| Gastroenteropankreaattinen endokriininen järjestelmä | |
| Enterosyytit | |
| biologiset nesteet | |
| Prosessit | |
Ruoansulatuskanavan motiliteetti | |